GMPPA-CDG Cure Odyssey
Using worm and fly avatars, we identified three hits with a shared pharmacophore that appears to target another CDG gene. We're testing a metabolic rebalancing hypothesis in patient fibroblasts next.


In collaboration with




Like most of the CDG drug repurposing projects we launched over the past year, we met the pioneer family in San Diego at the 2022 CDG CARE Family Conference sponsored by Sanford Burnham Prebys. The Roetting family is no exception. Their daughter Rosie is living with GMPPA-CDG, one of the rarest of the 150+ CDGs.
For those of you who are not professionally trained glycobiologists, the GMPPA gene encodes an enzyme called mannose-1-phosphate guanyltransferase alpha. The protein is a regulatory subunit of GMPPB, and the complex is responsible for the conversion of mannose-1-phosphate and GTP to GDP-mannose, a reaction involved in the initial synthesis of N-linked oligosaccharides. It’s an unusual but not entirely unheard of scenario in biology. GMPPA has lost its own catalytic ability in order to keep its fraternal twin GMMPB, and by extension GDP-mannose levels, in check.
Why does the cell care so much about its GDP-mannose levels? GDP-mannose is the rocket fuel for N-linked glycosylation, and as such the levels of GDP-mannose are finely tuned by the cell so as to maintain metabolic balance: not enough GDP-mannose results in too little glycosylation, but too much GDP-mannose leads to excessive glycosylation, which can be just as problematic.
GMPPA-CDG was first described in the medical literature as a new disease a decade ago by Koehler et al., 2013. In that study, a homozygous loss-of-function variant in GMPPA was confirmed by whole exome sequencing in three affected siblings from a consanguineous family. At first, the clinical team thought they were dealing with something called triple A syndrome because the affected children presented with a unique cluster of three clinical features: achalasia (difficulty or inability to swallow), alacrima (no tear production), and adrenal insufficiency. In their paper, Koehler et al showed that patient lymphocytes (immune cells) from the original GMPPA-CDG family have constitutively elevated levels of GDP-mannose compared to controls.

Of note: another CDG, NGLY1 deficiency, also presents with alacrima. GMPPA and NGLY1 deficiencies are both predicted to result in a net increase in glycosylation. In the case of NGLY1, it’s because glycan chains are not clipped off proteins. In the case of GMPPA, it’s because extra mannose residues — “hypermannosylation” — are incorporated into glycan chains. Specific proteins misbehave when their glycosylation status is altered in either direction, but we also know from other CDGs that, zooming out, metabolic flux is out of whack, too.
So what metabolic imbalance is caused by GMPPA deficiency? You can see in the figure below that GMPPA and its partner in crime GMPPB are the next station in the bucket brigade after PMM2, the enzyme that generates mannose-1-phosphate that gets converted to GDP-mannose by GMPPB. GDP-mannose then gets attached to dolichol phosphate in the first committed step of lipid-linked oligosaccharide (LLO) biosynthesis.

Based on our PMM2-CDG experience with epalrestat, we expect that a key rate-limiting enzyme located upstream or downstream of the primary enzyme deficiency is the dial that needs to be turned down — but not necessarily off — to restore metabolic balance. Perlara 1.0’s experiences with PMM2-CDG and NGLY1 deficiency are particularly instructive here. Worm, and fly models of deficient N-glycosylation synthesis all display slow growth, larval structural deficits, or lethality phenotypes. Slow growth can be rescued in a pathway-agnostic high-throughput chemical suppressor screen aka drug repurposing.
Although GMPPA is not characterized as having conservation as a regulatory subunit of GMPPB orthologs in other organisms, there is high homology of GMPPA to the worm gene Y247DA.1 and the fly gene CG8207. The GMPPA worm and fly orthologs are not as well characterized as in the case of PMM2-CDG, however there is remarkable conservation of these uncharacterized genes to the human GMPPA. The worm ortholog, Y47DA.1, is characterized as an NTP-transferase domain-containing protein and has conservation. The fly ortholog, CG8207, is loosely characterized as a nucleotidyltransferase.
Rosie is compound heterozygous for two missense mutations in GMPPA at positions either highly conserved at R318 or closely conserved as serine or threonine at T292. T292 in humans is S292 in both worm and fly, considered closely homologous due to the chemical similarity of serine and threonine. R318 is conserved in all animals, including human, worm, and fly.

Rosie’s variants are computationally predicted to be damaging but the exact defect or defects caused by each mutation – protein misfolding, protein instability, protein mis-localization, catalytic site inactivation, among others – has not yet been elucidated.
We turned to In Vivo Biosystems to generate worm avatars of Rosie’s two variants. Two independent clones were generated for each variant. A GMPPA knockout worm avatar was also generated for comparison. We suspected that the Rosie worm avatars experience protein misfolding stress due to excessive mannosylation. The simplest way to test for that is a heat shock assay, as shown below.

The R318W and T292P worm avatars have reduced activity at 37˚C. The GMPPA knockout behaves similarly. However, the Rosie worm avatar distributions overlap with the wildtype control (called N2). Nevertheless were were encouraged that we observed any differences. In search of a more robust disease phenotype to rescue, we next assessed developmental timing as expressed as body size over time.

The Rosie worm avatars as well as the knockout strain exhibit developmental delay at larval stage 1 and to a slightly lesser extent at larval stages 2 and 3. By larval stage 4 and Day 1 of adulthood, both the avatars and the knockout catch up to wildtype. The effect sizes observed in the heat shock and developmental timing assays are sufficient for hit validation studies but not large enough for a primary drug screen. So we subjected the Rosie worm avatars and knockout mutant to another stress test.
Recall that GMPPA and NGLY1 deficiencies both have alacrima as a clinical presentation. The two CDGs are also united by the fact that the metabolic scales are tipped toward excess glycosylation. For those reasons, we hypothesized that GMPPA worm avatars might be hypersensitive to the proteasome inhibitor bortezomib. Perlara 1.0 had success with using bortezomib as a sensitizer in the PMM2-CDG and NGLY1 deficiency worm drug repurposing projects. IVB did the experiment and the results are shown below.
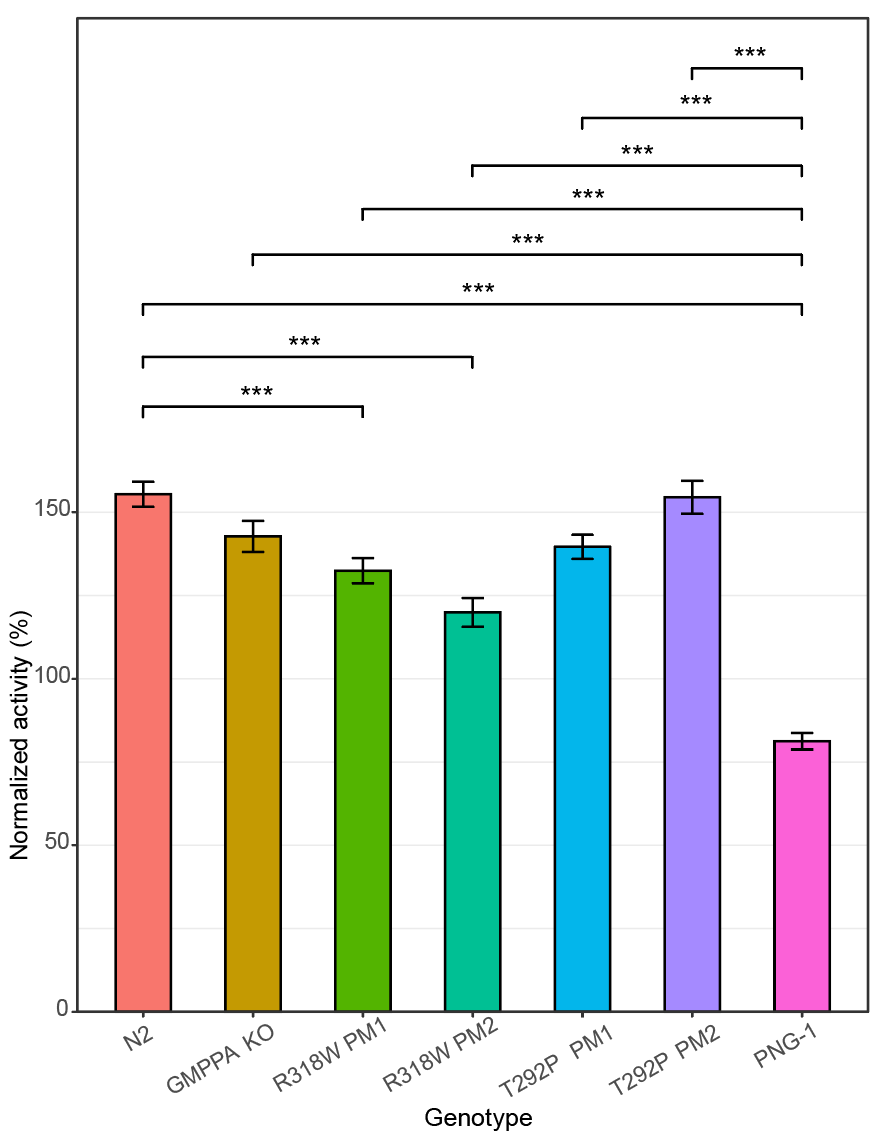
Bortezomib had the expected sensitizing effect on a NGLY1 knockout worm. However, compared to the NGLY1 knockout mutant, bortezomib did not have as strong an effect on the Rosie worm avatars or the GMPPA knockout mutant. The Rosie worm avatars would be appropriate for hit validation studies but the phenotypic differences were not robust enough for a primary drug screen. It’s hard to predict beforehand which disease models will pan out, which is why we advise our clients to construct a diversified portfolio of disease models.
Anticipating this outcome, we worked in parallel with Professor Clement Chow at the University of Utah to develop and characterize a GMPPA fly model for a drug repurposing screen.
The Chow lab is developing fly avatars for a variety of monogenic diseases, including CDGs, and then deploying said fly avatars in drug repurposing screens. For MAN1B1-CDG, a fly avatar was generated and screened, resulting in a group of hit compounds with the same mechanism of action. (A 1-to-N study has been underway since last Thanksgiving and we hope to say more about it soon).
Reminiscent of NGLY1 deficiency, GMPPA deficiency in flies also results in pupal lethality, i.e., animals die in the pupal case, as shown below:
So the readout for the screen is super simple: are there any living adults in the vial? ~1,500 compounds were screened by hand, a process that took several months from beginning to end. The screen yielded six hits. Upon retesting, 3/6 hits showed reproducible and robust rescue of pupal lethality. Remarkably, the top hit rescued pupal lethality to such an extent that 36% of the animals developed to adulthood — versus 0% in the negative control. What’s even cooler is the the top three hits are not only structurally related but also appear to target another CDG gene!
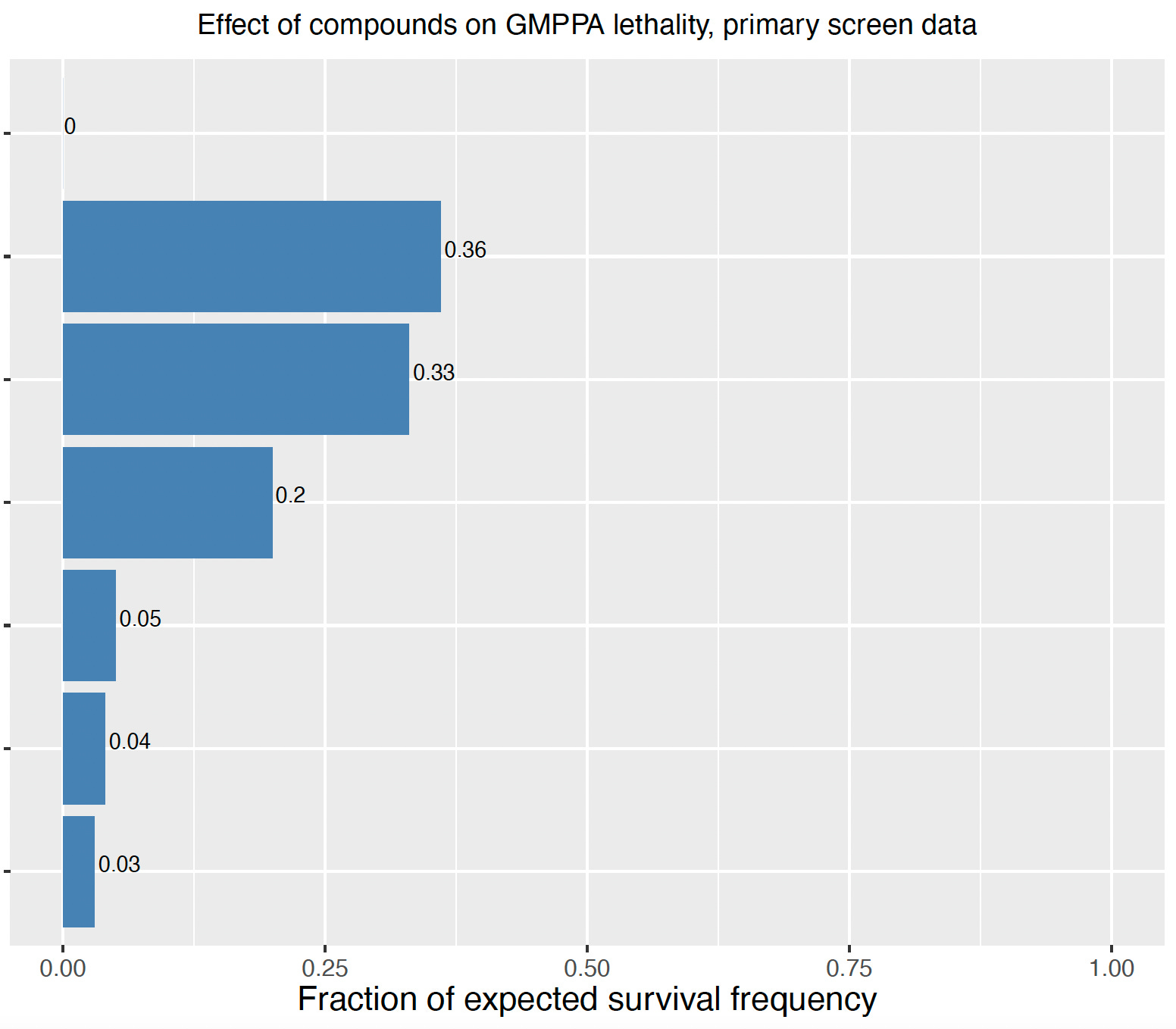
Worm hit validation studies with the six compounds were recently completed by IVB. The results for the top three hits show a trend toward rescue. Differences in solubility and stability in worm media, and in worms themselves, may explain why we don’t see perfect concordance between rescue in worms and rescue in flies. (Recall that in our NGLY1 worm and fly drug repurposing screens, only one compound was a hit in both species primary screens).
We’re planning on testing GDP-mannose levels in Rosie’s fibroblasts as well as three other GMPPA-CDG patients this summer. We also have a GMPPA KO cell line. As we’ve seen before in the cases of PMM2-CDG and SRD5A3-CDG, the mechanism of action of repurposing hits involves metabolic rebalancing.